Arabidopsis Epitranscriptome: Clock Gene Knockouts
Evening and midnight collections capture the Arabidopsis clock network at two late-day checkpoints, spanning wild type and single-gene knockouts in PRR and MYB/RVE clock modules.
| Field | Value |
|---|---|
| Organism | Arabidopsis thaliana (Col-0 background) |
| Collection times | CT20 (evening), CT24 (midnight) |
| Genotypes sampled | Col (WT), prr9, rve4, cca1, prr7 |
| Sample IDs (CT20) | Col_CT20, prr9_CT20, rve4_CT20 |
| Sample IDs (CT24) | cca1_CT24, prr7_CT24 |
CT = collection time (CT00 = midnight).
Samples and data provided by the Michael lab at Salk Institute.
Open IGV session (gene pileups)Introduction to the Plant Circadian Clock
Plants do not just respond to light and temperature in the moment. They also anticipate daily change. The circadian clock is an internal timing system that helps a plant align biology with the day–night cycle, so processes like metabolism, growth, and stress responses occur at appropriate times of day.
In Arabidopsis, this timing system is built from a set of clock genes that regulate one another in feedback loops. Together, they generate rhythmic activity that repeats about every 24 hours, even under constant conditions. One common way to visualize these rhythms is to fuse a clock-controlled promoter to a luciferase (LUC) reporter. As clock-regulated transcription rises and falls, luminescence oscillates in a regular wave over time (Panel A).
Importantly, the circadian system is not controlled only at the level of DNA and transcription. The cell also tunes clock behavior after an mRNA is made, through processes such as RNA processing and chemical modifications. One of the best-studied RNA marks is m6A RNA methylation, which is deposited by a dedicated writer complex and is enriched on transcripts from clock-regulated and clock-associated genes. In plants, m6A can help connect environmental light signals to clock regulation, for example through light-sensing cryptochromes that recruit m6A machinery to influence clock transcripts.
When individual clock genes are disrupted, the oscillator can keep time differently. Some mutations make the rhythm run faster (short period), some make it run slower (long period), and some weaken rhythmicity altogether (arrhythmic) (Panel B). These oscillator changes matter because the clock helps coordinate downstream "output" traits, including seedling growth (often measured as hypocotyl length) and seasonal development such as photoperiodic flowering (Panel C). Changes to RNA methylation and RNA processing can also shift clock timing, and reduced m6A activity has been associated with longer circadian periods in Arabidopsis.
This demo dataset provides late-day snapshots (CT20 and CT24) from wild type and selected clock-gene knockouts. It is designed to illustrate how perturbing the clock can be reflected in molecular readouts, including RNA-state and epitranscriptomic signatures, at defined times of day.
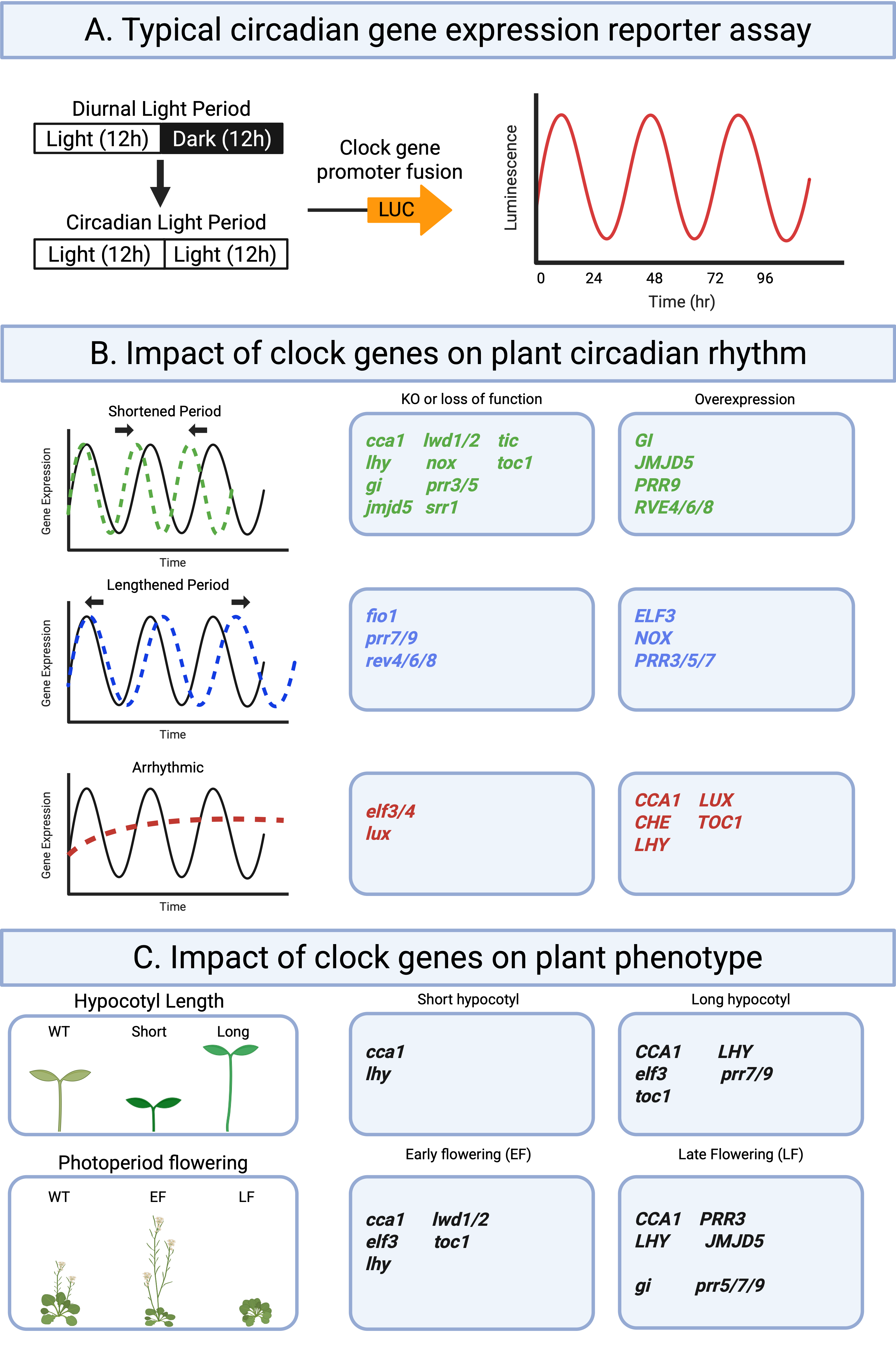
Panel A: Reporter assay logic (LUC luminescence rhythms)
Panel B: Oscillator phenotypes (short period, long period, arrhythmic) and example genes
Panel C: Output phenotypes (hypocotyl length; photoperiodic flowering) and example genes
Adapted from Current Biology.
CCA1
CIRCADIAN CLOCK ASSOCIATED 1
Clock role
CCA1 is a MYB-domain transcription factor embedded in the Arabidopsis core oscillator. In the canonical feedback architecture, CCA1 (with LHY) represses evening-phased components such as TOC1 via promoter binding.
Phenotypes
- Loss-of-function is associated with a short-period oscillator phenotype.
- Associated with short-hypocotyl output phenotypes.
- Associated with early-flowering output phenotypes.
Dataset sample(s)
cca1_CT24 (CT24); WT: Col_CT20 (CT20)
PRR7
PSEUDO-RESPONSE REGULATOR 7
Clock role
PRR7 participates in a morning-phased regulatory circuit: CCA1/LHY promote PRR7 expression, while PRR7 contributes repressive regulation that helps shape morning factor expression.
Phenotypes
- Loss-of-function is associated with a long-period oscillator phenotype.
- Associated with long-hypocotyl output phenotypes.
- Associated with late-flowering output phenotypes.
Dataset sample(s)
prr7_CT24 (CT24); WT: Col_CT20 (CT20)
PRR9
PSEUDO-RESPONSE REGULATOR 9
Clock role
PRR9 is part of the morning loop promoted by CCA1/LHY. The review describes PRR-family regulation as transcriptional repression that helps coregulate the morning oscillator factors.
Phenotypes
- Loss-of-function is associated with a long-period oscillator phenotype.
- Associated with long-hypocotyl output phenotypes.
- Associated with late-flowering output phenotypes.
Dataset sample(s)
prr9_CT20 (CT20); WT: Col_CT20 (CT20)
RVE4
REVEILLE 4
Clock role
RVE4 is a REVEILLE-family clock-linked regulator. RVE4 is a paralog of RVE8, which is highlighted in the reference figure as affecting circadian period; RVE4 is expected to behave similarly at the level of oscillator/output coupling.
Phenotypes
- REVEILLE-family perturbations (example: RVE8 in the figure) are associated with circadian period phenotypes.
- Use the figure's RVE8 example as context for expected oscillator-linked outputs.
- Do not add additional phenotype claims beyond what is shown for RVE8 in the figure.
Dataset sample(s)
rve4_CT20 (CT20); WT: Col_CT20 (CT20)
References
-
Complexity in the Wiring and Regulation of Plant Circadian Networks.
Current Biology
22 (2012) R648–R657.
(Corrected figure and erratum: Current Biology
23 (2013) 95–96.)
doi:10.1016/j.cub.2012.12.016
-
Current Insights into m6A RNA Methylation and Its Emerging Role in Plant Circadian Clock.
Plants
2023, 12, 624.
doi:10.3390/plants12030624