

Pseudouridine: Minor Isomer, Major Consequences for RNA Function
Pseudouridine: Minor Isomer, Major Consequences for RNA Function
Pseudouridine is the most abundant RNA modification and one of the most functionally versatile. It reshapes RNA structure, translation, splicing, and immune recognition, making it increasingly relevant to mRNA therapeutics, immunology, cancer biology, and translational epitranscriptomics.



RNA modifications as a regulatory layer
Cancer biology and immunology have largely been interpreted through DNA variation, transcription, and protein signaling. Those layers remain foundational, but they do not always explain phenotype alone. Tumors with similar genomic features can diverge in clinical behavior. Immune responses can shift rapidly without corresponding transcriptional changes. These gaps often reflect regulation occurring after transcription, in RNA processing, localization, turnover, and translation.
RNA modifications provide a chemical mechanism for this post-transcriptional control. Collectively termed epitranscriptomics, these marks are installed on RNA after transcription without changing the underlying sequence, yet they influence RNA stability, splicing, translation efficiency, and interactions with RNA-binding proteins and innate immune sensors.1 Among the more than 170 known RNA modifications, pseudouridine is the most abundant and arguably the most functionally diverse.
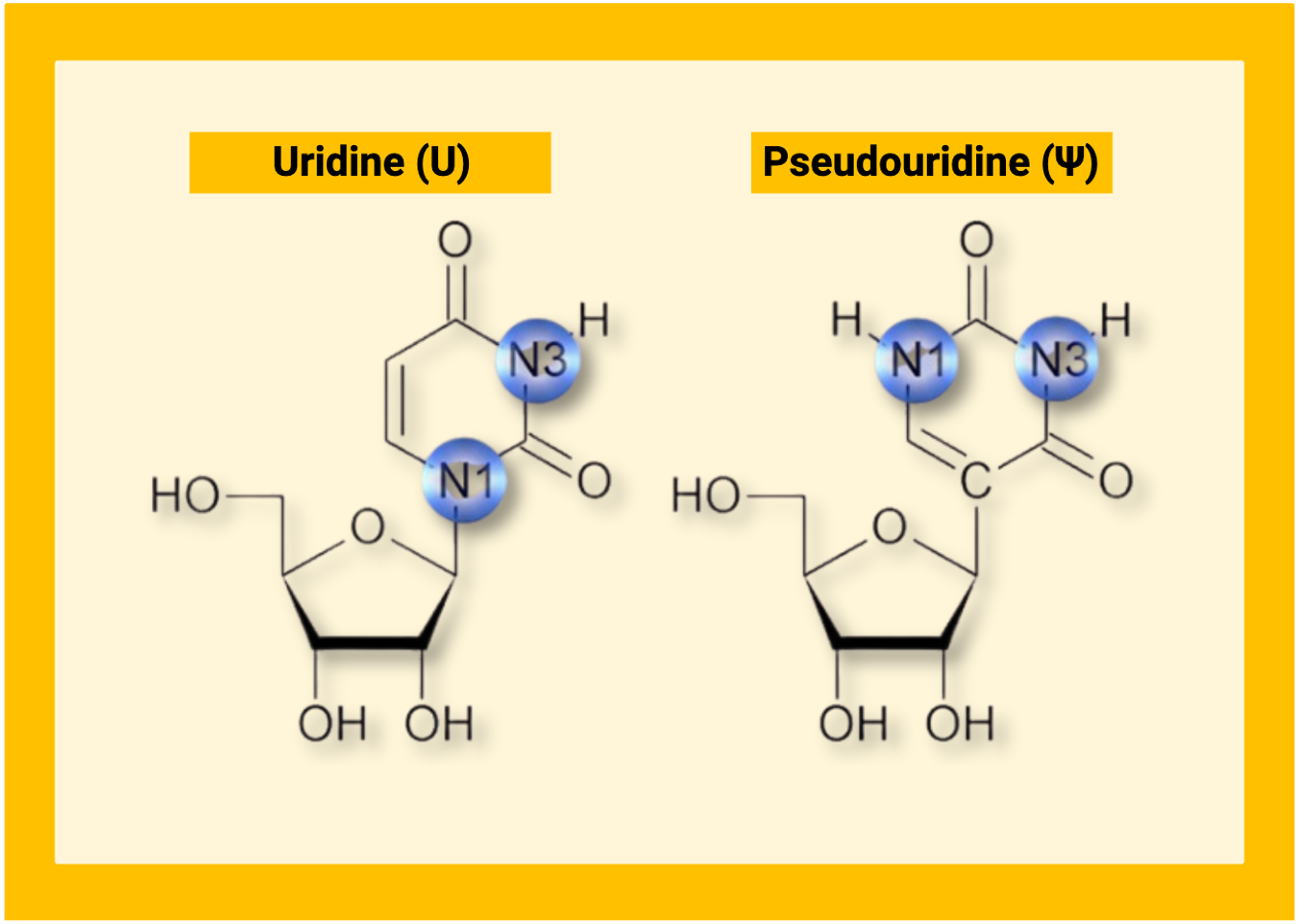
What is pseudouridine and how is it installed?
Pseudouridine (Ψ) is a structural isomer of uridine. The uracil base remains intact, but its connection to the ribose sugar is rearranged from a carbon-nitrogen bond to a carbon-carbon bond. This seemingly small change alters hydrogen bonding and stabilizes local RNA folding and stacking.2,3 The result is an RNA that behaves differently without reading differently on a standard sequencer, which makes pseudouridine both biologically important and technically challenging to study.
Pseudouridine is installed by over 10 distinct pseudouridine synthase (PUS) enzymes. Some act as stand-alone proteins that recognize local RNA structures. Others work through H/ACA ribonucleoprotein complexes that use guide RNAs to direct modification to specific sites.4 Pseudouridylation is not uniform across the transcriptome. It is shaped by enzyme specificity, RNA structure, and cell state.5 TRUB1, for example, is the predominant PUS enzyme acting on mammalian mRNA, recognizing conserved stem-loop features to direct site-specific modification.6
What pseudouridine does
Translation, decoding, and stop codon readthrough
Pseudouridine has long been central to translation through its presence in rRNA and tRNA. More recently, pseudouridylation within coding sequences of mRNA has been shown to alter translation directly, with effects that appear to depend on local sequence and structural context.7 One of the most striking functional demonstrations is stop codon readthrough. Targeted pseudouridylation can convert a premature stop codon into a sense codon, allowing ribosomes to read through what would otherwise be a disease-causing truncation signal.8 Recent work has continued to refine the practical parameters of this approach for therapeutic use.9
Splicing and RNA processing
Pseudouridine synthases modify human pre-mRNA co-transcriptionally, and there is good evidence that these events influence splicing outcomes.10 Changes in splicing shift isoform usage and protein domain composition, with downstream effects on interaction networks and cellular behavior. This provides a concrete mechanistic bridge between modification chemistry and pathway-level phenotype, particularly in the context of cellular stress and disease.11
Immune recognition
Innate immune sensors distinguish self from non-self RNA partly through nucleoside chemistry. Pseudouridine and related modified nucleosides suppress activation of Toll-like receptors and related sensing pathways.12 This property has turned out to have significant consequences for both disease biology and the design of therapeutic RNA.
Disease relevance and therapeutic applications
mRNA therapeutics and vaccines
The most recognized application of pseudouridine biology is in therapeutic RNA design. Incorporating pseudouridine into mRNA improves translational output and biological stability while reducing immunogenicity.13 N1-methylpseudouridine, a related modification, further enhances these properties.14 This chemistry is the molecular foundation of the mRNA vaccines used against SARS-CoV-2, and it is now driving a broader pipeline of mRNA therapeutics and personalized cancer vaccines in active clinical development.
Autoimmune diseases
The same pseudouridine properties that make immunogenic vaccines work can be redirected toward immune tolerance. Krienke et al. showed in preclinical MS models that pseudouridine-modified mRNA encoding myelin autoantigens can halt disease progression.15 The mechanism relies on the modification’s non-inflammatory character. Because the modified mRNA does not trigger innate immune co-stimulation, antigen is presented without the danger signals that would normally license effector T cell activation. The immune system instead generates regulatory T cells, which suppress the autoimmune attack on myelin without broadly shutting down immune function. This is an important distinction from conventional immunosuppressants, which carry real risks of infection and malignancy precisely because they blunt immunity systemically. Moderna’s mRNA-1195 has advanced this concept into Phase 2 trials for MS, and this technology is being explored for celiac disease and Type 1 diabetes as well. If the preclinical mechanism holds in humans, pseudouridine will have enabled two clinically meaningful RNA therapeutic strategies built on the same chemical modification, one that activates immunity and one that selectively quiets it.
Cancer and immuno-oncology
Pseudouridylation machinery is emerging as both a driver of tumor progression and a potential therapeutic target. PUS7 is overexpressed in glioblastoma relative to normal brain tissue, and high expression correlates with worse patient survival. Mechanistically, PUS7-mediated tRNA pseudouridylation promotes tumorigenesis, and small molecule inhibitors of PUS7 suppress tumor growth and extend survival in preclinical models.16 In a separate oncogenic context, MYC transcriptionally activates PUS7 to pseudouridylate the MCTS1 mRNA, enabling cancer cells to sustain proliferation under metabolic stress.17 Taken together, these findings suggest that PUS enzymes can be co-opted by oncogenic programs to reshape translational landscapes in ways that support tumor survival.
Pseudouridylation also connects to cancer immunity more directly. Targeting PUS1 activates antiviral innate immune signaling and enhances response to immunotherapy in preclinical cancer models,18 pointing toward potential combination strategies. The broader disease relevance of this machinery is further illustrated by dyskeratosis congenita, an X-linked syndrome caused by mutations in dyskerin, the catalytic component of H/ACA ribonucleoprotein complexes. The syndrome is characterized by accelerated aging phenotypes and strong cancer predisposition, a sobering reminder of how much depends on pseudouridylation machinery functioning normally.11,19,20
Emerging disease contexts
Pseudouridine-related biology is showing up across a widening range of disease areas, though the mechanistic evidence varies considerably in maturity. In women’s health, altered pseudouridylation has been linked to pregnancy-associated conditions including preeclampsia, where changes in RNA modification patterns may reflect broader shifts in RNA turnover and cellular stress responses during placental development.21 Neurodevelopmental contexts are also emerging, with pseudouridylation implicated in RNA regulatory processes relevant to brain development and disease, though causal mechanisms remain to be worked out in most cases.22 In some of these settings, pseudouridine measurements may reflect global RNA dynamics rather than a single targeted regulatory axis, which is an important interpretive caution as the field expands. Taken together, these connections suggest that pseudouridine biology is not confined to a narrow set of disease contexts but operates broadly wherever RNA processing and cell state intersect.
Why detection has been a bottleneck
Pseudouridine does not generate a simple, consistent signature during reverse transcription, which makes transcriptome-wide detection genuinely hard. Most methods have relied on chemical derivatization and computational modeling, each of which introduces biases and limits comparability across platforms.23,24 Recurring practical constraints include limited sample input, difficulty quantifying modification stoichiometry, the challenge of connecting a modified site to a functional consequence, and reproducibility across methods and studies.
Direct RNA sequencing via nanopore platforms offers a path toward profiling modifications in native RNA,25 though interpretation, reproducibility, and benchmarking are still being worked out. The field is moving, however deliberately, from asking whether a site is modified toward asking how much, in which cell states, and to what effect.
Closing perspective
RNA modifications are best understood as a state layer between expression and phenotype. Transcript abundance is often necessary context but rarely sufficient. Modifications can shift RNA stability, splicing, translation, and immune sensing in ways that change cell behavior without large changes in gene expression levels.
Pseudouridine occupies a distinctive position within this layer. It is structurally grounded, enzymatically regulated, stress-responsive, and directly connected to therapeutic applications that are already in the clinic. Its biology is most fully interpreted alongside complementary modifications. Pseudouridine reports on structural dynamics and translational control. m6A reports on RNA fate and decay. Inosine marks dsRNA surveillance and innate immune tone. These modifications reflect distinct regulatory axes, and measuring one while blind to the others can obscure the full picture.
New commercial solutions are addressing this gap by enabling the integrated, simultaneous profiling of pseudouridine, m6A, and inosine from a single sample. This allows researchers to move from single-mark observation to a more layered understanding of the epitranscriptomic landscape.
Explore additional posts in our Mods in Motion series
Blog Posts
- The Power of Inosine: How RNA Editing Shapes the Transcriptome
- A Beginner’s Guide to m6A and Other RNA Modifications
- Beyond Transcriptomics: RNA Modifications Shape Cancer Progression
Podcasts
About the author
James Tsay
James M. Tsay, PhD, is a biotech scientist and industry leader whose work spans across genomics, molecular diagnostics, and multi-omic technologies. He earned his PhD in physical chemistry at UCLA and completed postdoctoral research in biophysics at UC San Diego. He is currently a Scientific Advisor at AlidaBio, and previously held senior scientific and technical leadership roles at Illumina and Pleno Biosciences, contributing to the development of next-generation sequencing and advanced molecular profiling platforms.
References
- Roundtree, I.A., Evans, M.E., Pan, T., He, C. (2017). Dynamic RNA modifications in gene expression regulation. Cell, 169(7), 1187-1200. https://doi.org/10.1016/j.cell.2017.05.045
- Davis, D.R. (1995). Stabilization of RNA stacking by pseudouridine. Nucleic Acids Research, 23(24), 5020-5026. https://doi.org/10.1093/nar/23.24.5020
- Kierzek, E., Malgowska, M., Lisowiec, J., Turner, D.H., et al. (2013). The contribution of pseudouridine to stabilities and structure of RNAs. Nucleic Acids Research, 42(5), 3492-3501. https://doi.org/10.1093/nar/gkt1330
- Ge, J., Yu, Y.-T. (2013). RNA pseudouridylation: new insights into an old modification. Trends in Biochemical Sciences, 38(4), 210-218. https://doi.org/10.1016/j.tibs.2013.01.002
- Gilbert, W.V. (2024). Recent developments, opportunities, and challenges in the study of mRNA pseudouridylation. RNA, 30(5), 530-536. https://doi.org/10.1261/rna.079975.124
- Safra, M., Nir, R., Farouq, D., Vainberg Slutzkin, I., Schwartz, S. (2017). TRUB1 is the predominant pseudouridine synthase acting on mammalian mRNA via a conserved code. Genome Research, 27, 393-406. https://doi.org/10.1101/gr.207613.116
- Eyler, D.E., Franco, M.K., Batool, Z., Wu, M.Z., et al. (2019). Pseudouridinylation of mRNA coding sequences alters translation. PNAS, 116(46), 23068-23074. https://doi.org/10.1073/pnas.1821754116
- Karijolich, J., Yu, Y.-T. (2011). Converting nonsense codons into sense codons by targeted pseudouridylation. Nature, 474(7351), 395-398. https://doi.org/10.1038/nature10165
- Adachi, H., Pan, Y., He, X., Chen, J.L., et al. (2023). Targeted pseudouridylation: an approach for suppressing nonsense mutations in disease genes. Molecular Cell, 83(4), 637-651. https://doi.org/10.1016/j.molcel.2023.01.009
- Martinez, N.M., Su, A., Nussbacher, J.K., Schaening, C., et al. (2022). Pseudouridine synthases modify human pre-mRNA co-transcriptionally and pre-mRNA processing. Molecular Cell, 82(3), 645-659. https://doi.org/10.1016/j.molcel.2021.12.023
- Guillen-Angel, M., Roignant, J.Y. (2024). Exploring pseudouridylation: dysregulation in disease and therapeutic potential. Current Opinion in Genetics & Development, 87, 102210. https://doi.org/10.1016/j.gde.2024.102210
- Kariko, K., Buckstein, M., Ni, H., Weissman, D. (2005). Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification. Immunity, 23(2), 165-175. https://doi.org/10.1016/j.immuni.2005.06.008
- Kariko, K., Muramatsu, H., Welsh, F.A., Ludwig, J., et al. (2008). Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Molecular Therapy, 16(11), 1833-1840. https://doi.org/10.1038/mt.2008.200
- Andries, O., Mc Cafferty, S., De Smedt, S.C., Weiss, R., et al. (2015). N1-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA. Journal of Controlled Release, 217, 337-344. https://doi.org/10.1016/j.jconrel.2015.08.051
- Krienke, C., Kolb, L., Diken, E., Streuber, M., et al. (2021). A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science, 371(6525), 145-153. https://doi.org/10.1126/science.aay3638
- Cui, Q., Yin, K., Zhang, X., Ye, P., et al. (2021). Targeting PUS7 suppresses tRNA pseudouridylation and glioblastoma tumorigenesis. Nature Cancer, 2(9), 932-949. https://doi.org/10.1038/s43018-021-00238-0
- Ding, J., Bansal, M., Cao, Y., Ye, B., et al. (2024). MYC drives mRNA pseudouridylation to mitigate proliferation-induced cellular stress during cancer development. Cancer Research, 84(23), 4031-4046. https://doi.org/10.1158/0008-5472.CAN-24-1102
- Wang, F., Tong, Y., Guo, W., Qian, Y., et al. (2025). Pseudouridine synthase 1-targeted therapy activates antiviral immunity to boost cancer immunotherapy. Cell Reports, 44(9), 116233. https://doi.org/10.1016/j.celrep.2025.116233
- Bellodi, C., Kopmar, N., Ruggero, D. (2010). Deregulation of oncogene-induced senescence and p53 translational control in X-linked dyskeratosis congenita. EMBO Journal, 29(11), 1865-1876. https://doi.org/10.1038/emboj.2010.83
- Angrisani, A., Vicidomini, R., Turano, M., Furia, M. (2014). Human dyskerin: beyond telomeres. Biological Chemistry, 395(6), 593-610. https://doi.org/10.1515/hsz-2013-0287
- Jia, S., Yu, X., Deng, N., Gao, Z., et al. (2025). Deciphering the pseudouridine nucleobase modification in human diseases: From molecular mechanisms to clinical perspectives. Clinical and Translational Medicine, 15, e70190. https://doi.org/10.1002/ctm2.70190
- Yang, Q., Al-Hendy, A., Boyer, T.G. (2026). Molecular insights into widespread pseudouridine RNA modifications: implications for women’s health and disease. Biology, 15(2), 142. https://doi.org/10.3390/biology15020142
- Li, X., Zhu, P., Ma, S., Song, J., et al. (2015). Chemical pulldown reveals dynamic pseudouridylation of the mammalian transcriptome. Nature Chemical Biology, 11(8), 592-597. https://doi.org/10.1038/nchembio.1836
- Motorin, Y., Helm, M. (2019). Methods for RNA modification mapping using deep sequencing. Genes, 10(1), 35. https://doi.org/10.3390/genes10010035
- Begik, O., Lucas, M.C., Pryszcz, L.P., Ramirez, J.M., et al. (2021). Quantitative profiling of pseudouridylation using nanopore sequencing. Nature Biotechnology, 39(10), 1278-1291. https://doi.org/10.1038/s41587-021-00915-6
Supporting information
- Additional clinical trial context: Moderna mRNA-1195 multiple sclerosis relapse prevention study, ClinicalTrials.gov identifier NCT06735248.